
 One of the recurring discussions in evolutionary biology is the question of how to test adaptive hypotheses. Creationists often claim (wrongly) that the theory of natural selection is tautological, and so did, at some point, prominent philosopher of science Karl Popper (he later acknowledged that his comment was based on a misunderstanding of the theory, something that creationists strangely neglect to mention whenever they quote him in their favor — see this short essay I wrote about it for Skeptical Inquirer). And then, of course, there is the never ending issue of the quasi-scientific status of evolutionary psychology, precisely because adaptive hypotheses are particularly hard (though not impossible) to test in the case of Homo sapiens (more on this below).
One of the recurring discussions in evolutionary biology is the question of how to test adaptive hypotheses. Creationists often claim (wrongly) that the theory of natural selection is tautological, and so did, at some point, prominent philosopher of science Karl Popper (he later acknowledged that his comment was based on a misunderstanding of the theory, something that creationists strangely neglect to mention whenever they quote him in their favor — see this short essay I wrote about it for Skeptical Inquirer). And then, of course, there is the never ending issue of the quasi-scientific status of evolutionary psychology, precisely because adaptive hypotheses are particularly hard (though not impossible) to test in the case of Homo sapiens (more on this below).
It was therefore with delight that I recently happen to stumble on a paper by Mark Olson and Alfonso Arroyo-Santos entitled “How to study adaptation and why to do it that way: adaptation, just so stories, and circularity,” published in the prestigious Quarterly Review of Biology (pdf freely available here).
The basic argument put forth by Olson and Arroyo-Santos is that “circularity” comes in degrees, from vicious to virtuous, which is why they prefer the term “loopiness,” as in feedback loops between hypotheses and empirical evidence. Perhaps the best way to get what they are saying is to examine a few of their figures.
This one is a summary of the standard adaptationist story about why giraffes have long necks:

If you start with the observed pattern (lower portion of the figure), you will agree that, observationally, modern giraffes have long necks and they feed from tall trees. Based on that, you infer (upper portion of the figure) that the following things were true of the ancestors of modern giraffes: (i) they varied in neck length; (ii) this variation was heritable; (iii) short-neck giraffes had less access to food (functional argument); and (iv) short-neck giraffes, as a consequence of their phenotype, had lower fitness (adaptive argument).
The problem is, up to this point you have absolutely no evidence for any of inferences (i)-(iv) above. Which truly does make your “explanation” a just-so story.
But wait!, say — correctly — Olson and Arroyo-Santos, this can be the beginning of a fecund research program, because you can turn (i)-(iv) into hypotheses to be directly or indirectly tested.
For instance, evidence for (i) may be found in the fossil record; evidence for (ii) in extant populations (which would license the inference, though obviously not prove, that this was the case also in the past); (iii) may be tested directly in extant populations, or indirectly via optimization models; and (iv) is a reasonable conclusion from (i)-(iii), and therefore acceptable in direct proportion to the strength of the evidence in favor of (i)-(iii).
If you ponder the above for a minute you will realize why this shift from vicious circularity to virtuous loopiness is particularly hard to come by in the case of our species, and therefore why evolutionary psychology is, in my book, a quasi-science. Most human behaviors of interest to evolutionary psychologists do not leave fossil records (i); we can estimate their heritability (ii) in only what is called the “broad” sense, but the “narrow” one would be better (see here); while it is possible to link human behaviors with fitness in a modern environment (iii), the point is often made that our ancestral environment, both physical and especially social, was radically different from the current one (which is not the case for giraffes and lots of other organisms); therefore to make inferences about adaptation (iv) is to, say the least, problematic. Evopsych has a tendency to get stuck near the vicious circularity end of Olson and Arroyo-Santos’ continuum.
Here, by contrast, is an example of what virtuous loopiness looks like, in the case of the observation that swimming animals tend to have streamlined bodies, regardless of their taxon and phylogenetic relatedness:
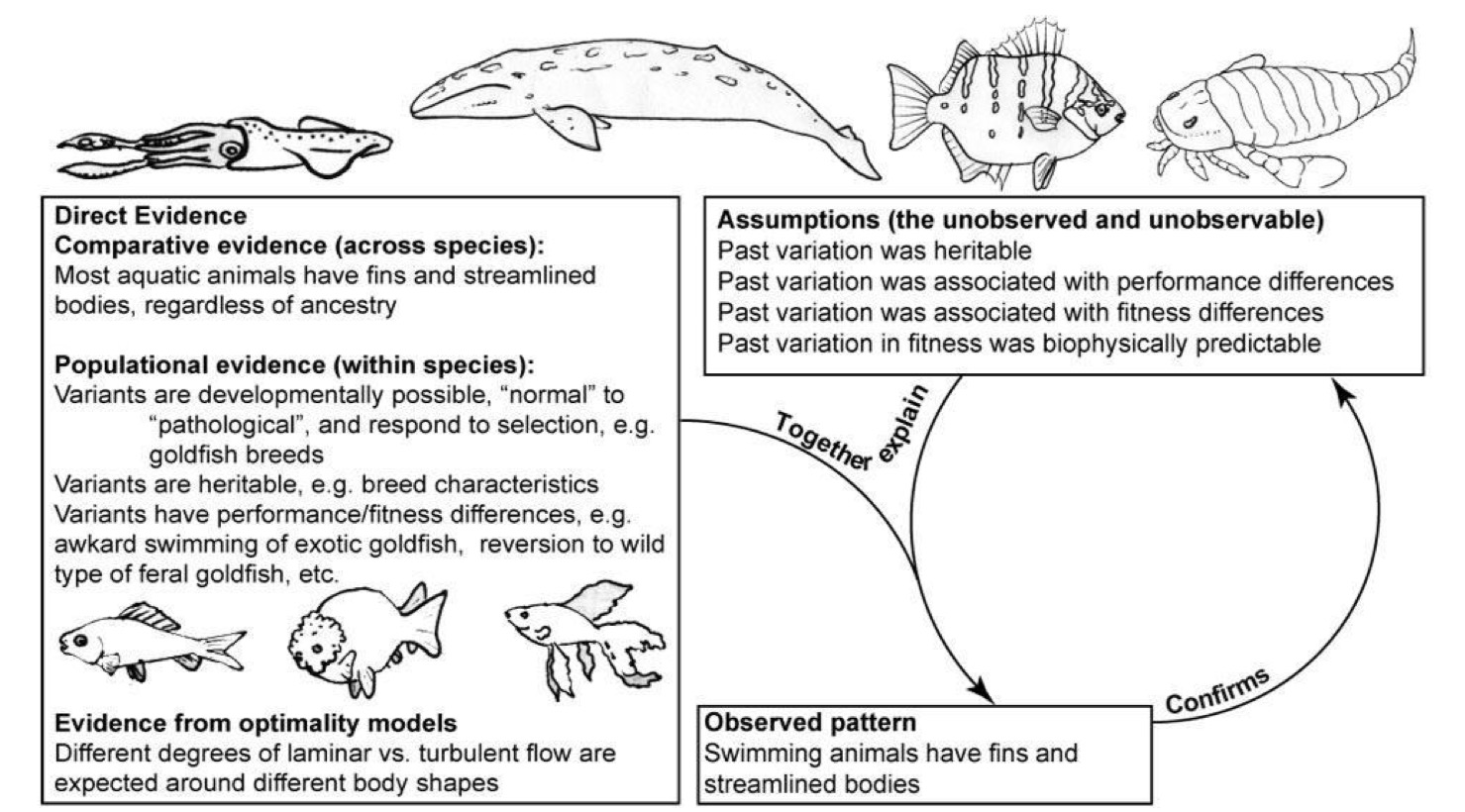
You can appreciate the much more clear distinction between the initial observation of a pattern (lower right), the development of a number of assumptions necessary to explain the pattern (upper right), and three types of empirical evidence that feed into the other two components (left side of the figure).
Let us take a closer look at that last bit. Olson and Arroyo-Santos argue, again, correctly, that there are fundamentally three types of evidence that bear upon any adaptive hypothesis: from the comparative method, from population biology, and from optimality considerations.
The comparative method has to do with contrasting large number of species with different degrees of phylogenetic relatedness. The methodology has been fine tuned decades ago, including the development of sophisticated statistical approaches and the use of computer simulations to assess how well different methods of phylogenetic reconstruction work. This is another area where evopsych suffers, however, since there are very few and widely scattered “close” relatives of our species (no other Homo surviving, a couple of species of chimpanzees and the gorillas, separated from us by several million years of independent evolution).
Population biology is a very broad field at the interface among ecology, evolutionary biology and genetics, again characterized by well honed methods that rely on basic population and quantitative genetic theory. This is where our estimates of population variation (and hence also heritability) come from.
Optimality modeling has to do with applying biomechanical considerations to the study of adaptation, developing quantitative assessments of how well certain biological structures — and their variants — perform their function in response to pertinent environmental stimuli. In the case of the streamlined bodies of fish, marine mammals, and some invertebrates like squids, the pertinent theory comes from hydrodynamics.
The third figure, also from the Olson and Arroyo-Santos paper, explains how the loopiness of adaptive explanations works in the recurring case of discussions about the relative roles of natural selection and developmental constraints:

These two classes of explanations of biological structures are too often, even by biologists, considered to be mutually exclusive, and indeed sort of divide evolutionary biologists from developmental biologists, as well as “externalists” (who favor external causes, like natural selection) from “internalists” (also known as structuralists, who favor internal causes, like developmental constraints).
But Olson and Arroyo-Santos rightly point out that this is yet another case of loopiness: in the figure, they show a typical scattergram that hints at a strong linear correlation between two traits, X and Y (say, body size and antler size in the famous “Irish elk,” see figure at top of post). The typical question is: why do the data points line out that way within the phenotypic space defined by the two variables? Why the empty spaces of unrealized phenotypes (say, of large bodied elks with small antlers, or small bodied ones with very large antlers)?
Each of the two alternative explanations, that the empty spaces are explained by the fact that some morphologies are maladaptive, or that they are developmentally inaccessible (i.e., there is no way for the organism in question to build that phenotype, given its genetic-developmental matrix) are testable by stretching the initial circularity into the same sort of loopiness we’ve seen before. Moreover, the two are not mutually exclusive, since developmental constraints can limit, but not nullify the action of natural selection, and selection in turn may favor a genetic-developmental matrix that precludes the accidental invasion of maladaptive areas of phenotypic space. There are, in fact, many examples in the literature of this sort of analysis, and they represent some of the best achievements of modern evolutionary biology.
Here is how Olson and Arroyo-Santos then conclude their excellent paper:
“Studies of adaptation would necessarily seem to require the sort of loopy reasoning [presented in this paper]. Recognizing how adaptationist explanations are structured in actual practice helps give clarity to problems that have plagued biology, such as debates over tautology/circularity, and resolve false conflicts, such as the mutual scorn that often characterizes the adherents of the comparative, population/quantitative genetics, and optimality approaches. Instead, as providers of complementary sources of direct evidence, no single approach has a monopoly on tests of adaptation. An understanding of the real, loopy structure of evolutionary explanation encourages biologists to discuss truly substantial issues awaiting attention, such as how to identify the population of hypotheses from which to select the ‘best’ explanation, how scientists know the best explanation when they see it, or how best to weave disparate sources of evidence into a single explanation. By accepting that studies of adaptation require multiples types of direct evidence, evolutionary biologists can improve current research practice by designing a compelling and long-overdue integration of comparative, populational, and optimality approaches.”

Massimo:
And language? Presumably you also think that the study of the evolution of language is a lost enterprise.
LikeLike
Hi Massimo,
This broad area has been called several different things (“ethology” and “sociobiology” in addition to those two). So long as we accept that we can in principle identify at least some modern human psychological traits as adaptive, and thus as related to our evolutionary past, then I’m happy with that (whatever the labelling).
Robin,
If what I’m trying to do is establish the principle (that we can sometimes relate psychological traits to our evolutionary past) then pointing to examples that are obvious to an 11-yr-old would seem a good way of doing it.
LikeLiked by 1 person
On the article:
Much of the article is agreeable. Under the causal closure assumption of methodological naturalism, it logically follows that all aspects of human behaviour (social and psychological) shall be explicable in naturalistic terms, and therefore, the only question that remains is, whether evolutionary psychology is that naturalistic science which can satisfactorily explain human behaviour.
I think there are two aspects to the issue: human behaviour as a result solely of evolutionary processes; and human behaviour as resulting from both evolutionary and extra-evolutionary (but naturalistic) processes. It is true, trivially and perhaps circularly, that the entire gamut of our behaviour is explicable in evopsycho terms, in that we will not be having any of these behaviours had evolution not bring it about. But the really interesting issue, as I see it, is emergentist: whether such behaviours (rape, altruism, love of art, suicide, etc) are causally entailed by natural selection, or whether nature selects certain traits by reasons other than fitness out of a list of other possible traits.
That is, supposing there is a behavioural trait x, and another trait y, and both these have more or less equal fitness values, and both these are not developmentally constraint. If in such circumstance it so happens that we develop triat x but not y, it will mean evopsycho is irrelevant, since adaptability is not the issue here; and therefore, evopsycho does not add to our understanding of why y occurs. In such circumstance it follows that other extra-evolutionary factors at play. And I’d say this is the likelier possibility for most of our behaviour.
For example, our decision to use the male’s surname instead of the female’s, may simply be a logical consequence of patriarchy, and while the dominance of patriarchy maybe accountable in evopsycho terms, the dominance of male surnames may simply be considered a logical consequence rather than an evolutionary one. Or, to take another example. Our prehistoric decision to practice animal sacrifices rather than plant sacrifices may not be explicable in evopsycho terms in that both practices have the same fitness consequences; therefore, the eventual predominance of animal (rather than plant, fungi, etc) sacrifices requires non-evolutionary explanations.
LikeLike
||
I love The Open Society and its Enemies. And important work, I would maintain, and right in many ways though obviously, not all.
||
As do I. Something we finally agree on!
I’d add that, if Popperian demarcation is thrown off the table, I’m not really sure whether there is another good methodological criterion for demarcating science from non-science. Falsification seems the best bet in town. The other alternative is Quinean webs, which with its underdetermination undertones may not be too palatable to the scientific realist however.
LikeLiked by 1 person
David,
Yes, genomic adaptations can be studied in humans, although there are a number of caveats there too. The major one is that the so-called genotype-phenotype mapping function is anything but linear and straightforward, so the genomic level only tells you so much abou the phenotypic one.
Bunsen,
Yeah, the evolution of language falls pretty much under the evolution of cognition umbrella. Again, no clue as to the selective pressures, obviously no fossil record (except for the fact that we known approximately when it happened, based on the anatomy of the voice box, which does preserve), and no meaningful inter-species comparisons.
Coel,
“This broad area has been called several different things (“ethology” and “sociobiology” in addition to those two)”
No, you are mixing a bunch of different things. Ethology is the general study of animal behavior, and it doesn’t necessarily have an evolutionary bent. Sociobiology was the precursor of evopsych, and suffered from the same problems.
It really comes down to the fact that if evopsychs simply want to do human behavioral ecology then they are not doing anything new, and their work is going to be difficult. If they want to establish a new field focused on evolutionary interpretations of modern behaviors and based on the idea of massive brain modularity then they are pretty much hopeless.
Haulianlal,
Seems to me you are essentially talking about the role of cultural evolution, and you are right. Unfortunately, we currently don’t have a good theory of cultural evolution (though plenty of people are working on it), and we don’t know how it integrates with biological evolution. And yes, understanding things like why many (not all!) human cultures pass on male surnames in terms of patriarchy is much more meaningful than inventing selection stories based on what may have been going on in the Pleistocene (when people presumably did not have surnames to begin with).
As for Popperian demarcation, I’ve argued in The Philosophy of Pseudoscience (http://tinyurl.com/kuxyjz7) that it definitely doesn’t work, but that there are more nuanced ways of doing demarcation.
LikeLiked by 1 person
Karl Popper was a co-founder of the Mont Pelerin Society. His Open Society was his first major publication in English. He most certainly saw a connection between his philosophy of science and his politics, and I don’t understand how anyone can just declare there isn’t one. I think there’s a (heavy) burden of proof to meet.
Philosophy of science bears directly on political practice. What after all is the scientific status of the claims of economics departments and policy institutes? Your colleagues at Bleeding Heart Libertarians know very well how scientificity connects to political philosophy, but you have no argument with them. I think their philosophy of science was designed to promote certain political goals. A radical epistemological skepticism that leaves their claims uncontested serves the same ends, does it not?
As for loving The Open Society…among its many failings is the extraordinary way Popper does not actually trouble to address other people’s ideas, but his notion of their ideas. And there is an equally persistent reliance on uncritical notions about the world and history, insofar as it deigns to engage the real world at all. There are other problems even more tightly connected with his philosophy of science. But his tendency to ignore facts is usually just called “lying.” Historically, religion and philosophy are inextricably intertwined, so I suppose this is appropriate.
LikeLike
Hi Massimo,
Agreed on both counts, though for someone like me who always wants to emphasize the continuity between different fields of knowledge the “newness” of a field is not that important.
Indeed the basics of evo psych can be traced back to Darwin, who in the OofS said: “In the distant future I see open fields for far more important researches. Psychology will be based on a new foundation, that of the necessary acquirement of each mental power and capacity by gradation”. Then in Descent of Man he gave much more consideration to how our psychological traits might have evolved.
I tend to think that the “modularity” hypothesis gets misunderstood. Modularity is from a functional perspective (in which case it can hardly be false, since what is selected for are behaviours), it is not a thesis about hardware (that thesis being obviously false). (And nothing in the modularity hypothesis says that modules cannot share information and cross-influence each other.)
LikeLike
A psychologist friend once implied that my theory below may display too much of an “evolutionary psychology” perspective. Conversely I expected him to tell me that it was already an accepted understanding! Up or down I’d appreciate any thoughts on it.
One useful way to distinguish the human from our humanoid ancestors, I think, is formal language. Though helpful for communication I consider language to mainly be important as a separate variety of thought itself. The monkey, the dog, and the modern human feral child do not have the capacity to think in this advanced way.
Another thing that seems unique about us is our amazingly expressive faces — you’ll not find this anywhere else in the animal kingdom. I propose that our unique formal languages brought the need for this unique facial expressiveness. Here is my theory:
By expressing the language form of thought it was now far more possible for us to deceive each other in personally beneficial ways. Though language gave us tremendous new abilities, the general deception that it fostered was also problematic — people naturally had an interest in saying things that contradicted how they felt, so in practice early humans couldn’t trust what they were telling each other. I suspect that one of evolution’s responses to this was to develop a vast array of muscles that involuntarily displayed extremely complex human emotions through the face. This would have given the listener some true information about the speaker’s mentality, and even seems important today when a good “poker face” can overcome these involuntary signs of how we feel.
(Wtc, I missed the deadline to respond to your final weekend comment, but you can find it here on my mostly untended site: https://physicalethics.wordpress.com/home-page-with-a-cocktail-party/a-final-authors-note/comment-page-1/#comment-117).
LikeLiked by 1 person
More on evolutionary psychology:
I think the proto-science is potentially useful for explaining behavioural and cultural traits that are more or less universal across societies. For example, it may one day satisfactorily explain why almost everyone have an eerie feeling of the dark, why ghost stories are invented across all cultures, why rape is frowned upon in virtually all societies, why we develop facial expressions to express emotions, why we cry as babies, why we develop laughter and smile as opposed to other forms of expressing ourselves, and so on. These are cases that are almost so universal that the exceptions prove the rule, and we may expect a science to pop out of these speculations someday.
Those traits that are culture-specific and are unique, need not appeal to evolutionary explanations however (which will make as much sense as appealing to gravity to explain why a World War II German plane was shot down over London skies). These, imo, require extra-evolutionary explanations by taking on the resources available from anthropology, history, non-evolutionary psychology, and so on.
Hi Massimo:
|| As for Popperian demarcation, I’ve argued in The Philosophy of Pseudoscience (http://tinyurl.com/kuxyjz7) that it definitely doesn’t work, but that there are more nuanced ways of doing demarcation. ||
I hav read your some chapters of your book, and some of your articles, in addition to other writers. My view is that while such nuanced accounts satisfy our drive to describe the nature of science, they do not work as prescriptions; and because they do not work as prescriptions, they are essentially useless for the demarcation problem.
While I accept your point that any sound philosophy of science must at least agree on the whole gamut of conventional science accepted by the vast community of scientists, the account fails exactly where its needed most: that is, how to adjudicate on fringe areas of the sciences.
Steve:
|| He most certainly saw a connection between his philosophy of science and his politics, and I don’t understand how anyone can just declare there isn’t one. ||
I concur. Fallibilism is a common thread connecting his philosophy of science and political philosophy, and I think this is rather important, even if it is a monumental failure. His insistence on the open society is essential for any epistemic investigation for the simple reason that, were a society not open, and each investigation is dictated by some political authority, there will be no way to correct errors; indeed, there will be no way to even know what is erroneous and what not.
Further, the free spirit inherent in everyone of us demands that we are essentially rebellious, at least in thought if not in deed or even word. Therefore, on humanitarian grounds, it is important that a political society is always open – even if we presume that the society is built on the most rational foundations, and we reach an utopian state of affairs. I also concur with Popper that liberty must be protected by force, by coercing those who preach ideas contrary to liberty (although this is again an open case).
As for your disdain of his philosophy, I really do not understand why, or how. Yes, he did misinterpret many thinkers, but Popper the exegete can be separated from Popper the philosopher.
LikeLike
Coel,
“for someone like me who always wants to emphasize the continuity between different fields of knowledge the “newness” of a field is not that important.”
But someone like you may underestimate (in this case, I think, by a lot) that there are qualitative differences out there. This isn’t a question of newness, but of serious methodological problems.
“Indeed the basics of evo psych can be traced back to Darwin”
Even the great ones get some things wrong. Again,it isn’t a question of the very idea of evopsych being incoherent or problematic. It’s the practice of it, and for reasons that have to do with the special phylogenetic and cultural-evolutionary properties of our species.
Haulianlal,
“My view is that while such nuanced accounts satisfy our drive to describe the nature of science, they do not work as prescriptions; and because they do not work as prescriptions, they are essentially useless for the demarcation problem.”
They are meant as descriptions. Useless? So I guess my map of the New York City subway is useless because it is descriptive rather than prescriptive?
LikeLike
It seems like one thing we humans would like to avoid, but never can – is making value judgments. This is true for distinguishing between science and pseudoscience – the gray areas are always going to require an evaluation of the methods employed. Short cuts are likely to lead one astray.
LikeLike
Massimo: “Seems to me you are essentially talking about the role of cultural evolution, and you are right. Unfortunately, we currently don’t have a good theory of cultural evolution (though plenty of people are working on it), and we don’t know how it integrates with biological evolution. “
To add to the problem, a lot of writers, in discussing human evolution, tend to blur the distinction between cultural and biological evolution.
LikeLike
Wtc,
Well, blurring the distinction between biological and cultural evolution is kind of inevitable, since there is no easily definable boundary between the two.
LikeLiked by 1 person
stevenjohnson: As I said, I quite like Open Society. That your political orientation is different from Popper’s — or mine — doesn’t mean that ours is wrong or that yours is correct or based on “facts” or better readings of historical figures.
Everyone thinks the other guy is being “uncritical” and that they themselves are paragons of critical reason. That’s the oldest song in the book.
LikeLiked by 2 people
Massimo: “Well, blurring the distinction between biological and cultural evolution is kind of inevitable, since there is no easily definable boundary between the two.”
The distinction I had in mind was between a million or so years of biological evolution (from other pre-humans) vs. less than 10,000 years from the beginnings of civilization, not nearly enough time for significant biological changes. The biological stuff is ours to keep, as long as Homo sapiens lasts; the cultural could disappear in a matter of hours, given a worst-case scenario. We tend to take this for granted as our birthright, without realizing how fragile it is, and what a mass of things every individual needs to learn and assimilate to become a full human being (shades of Pinocchio!). And someone like Noah Harari says we’re going to “evolve” into superhumans, or be replaced by the machines (but I enjoyed his first book). This is what I took from “The Open Society,” a frankly polemic book of the best kind, written in the darkest part of WW2, by an Austrian Jew in exile, looking at his homeland caught between Hitler and Stalin, one of the most positive and encouraging books I’ve ever read (sorry, I’ve got to put in a word for Popper, although he certainly needs no defense).
LikeLiked by 2 people
Hi Coel,
The point I am making is that in order for evpsych to be a viable field being a specialisation on its own, there has to be something in between “immediately obvious, even to an 11 year old” and “your guess is as good as mine”. It is not clear to me that there is much if anything at all in that gap.
LikeLike
Hi Massimo:
||So I guess my map of the New York City subway is useless because it is descriptive rather than prescriptive?||
Geographical maps do not just describe places but tells you where to go, and where not to, in order to reach a certain destination. Your account, and all other philosophies of science barring Popper’s and the Vienna Circle’s, do not have prescriptive content, therefore they cannot help in resolving the demarcation problem, or adjudicating the scientific or non-scientific status of a proposed theory. If it works as prescriptions, I do not see how.
Hi Michael:
||It seems like one thing we humans would like to avoid, but never can – is making value judgments. This is true for distinguishing between science and pseudoscience – the gray areas are always going to require an evaluation of the methods employed.||
Exactly. Any attempt to demarcate science from non-science cannot but be prescriptive. If it describes the nature of science, it must also tell us what science is not. The criterion should be simple in order to have prescriptive teeth. Otherwise, if it makes exceptions here and there for this or that established scientific theory, then the fields one normally considers to be pseudoscientific, can also be pushed in through the doors of such exceptions.
Hi wtc:
||The distinction I had in mind was between a million or so years of biological evolution (from other pre-humans) vs. less than 10,000 years from the beginnings of civilization, not nearly enough time for significant biological changes. ||
I think this is why evopsycho is useless to account for our specific behavioral and psychological attitudes, precisely because these attitudes develop ever so recently they cannot be the result of evolutionary processes.
I like your comment about Popper too. And I’d add further that if his demarcation criterion is a failure, it is a monumental failure and a great shame, because it is the best tool we have to not only demarcate science from pseudo science, but to ensure an open society based upon an epistemology (in that sense that nobody should attempt large-scale social engineering precisely because there is no way to tell if such endeavours are “correct” or “true”).
LikeLiked by 1 person
Robin,
First, isn’t it highly improbable that in any field there would be little or nothing between “blindingly obvious” and “pure guesswork”? That would be highly peculiar about any topic.
Second, there does seem to me sufficient of substance that comes under the broad banner “evo psych” to make it a valid field. If you look at something like Buss’s textbook “Evolutionary Psychology” (Fifth edn) there is much that would fall in your middle ground. (Which is not the same thing as accepting every claim ever made in the field.)
Even Massimo sees some valid stuff in some parts of evo psych (from a reply to a post by Jerry Coyne, where he disagrees on other points).
“To get us started, let’s look at some of the entries in Jerry’s list of recent successes (or at least examples of progress) in evopsych. Some are obvious and hard to dispute: incest avoidance, innate fear of dangerous animals, parent-offspring conflicts, and the like. As Kaplan and I (and plenty of others) have pointed out, these are the areas where evopsych is at its strongest because the target behaviors are common among mammals, or at least primates.”
Again, I really do think one has to evaluate each aspect of evopsych on its merits, rather than take a blanket dismissal.
One also has to be careful about dismissing evopsych by definition: “if it’s sound methodology and supported by evidence then, ipso facto, it’s not “evo psych”, it’s something else such as behavioral ecology; it only counts as evo psych if the method is ropey and the evidence sparse”.
The differences between these labels anyhow seem to me to be minor and arbitrary; the whole point of evopsych is to try to link things together, such as linking psychology with evolutionary biology. And if one says it’s ok to do this where you have animal correlates but not for uniquely human traits, then ok but the whole point of evopsych is to view humans as an animal and look for correlates in other animals. No-one says that an evolutionary account of the origin of the elephant nose is not do-able just because their noses are fairly unique.
LikeLike
Haulianlal,
Your disanalogy does not work: just like I have to follow the subway map IF I want to get somewhere (but I don’t have to, do I?), I have to pay attention to the descriptively demarcated science/pseudoscience territory IF I’d rather do science than pseudoscience.
LikeLike
Hi Coel,
There was a field called “Memetics” once and a Journal of Memetics. Pretty much filled the bill.
Incidentally – “fear of dangerous animals” would seem to fall into the “an 11 year old could work it out for herself” category.
Incest avoidance. Now I wonder if the scientific answer to this is really much better than the layman’s guess. There are two possibilities I can think of , or a mixture of the two.
I am pretty sure I didn’t say that, or anything like it.
LikeLike
Coel,
“One also has to be careful about dismissing evopsych by definition: “if it’s sound methodology and supported by evidence then, ipso facto, it’s not “evo psych”, it’s something else such as behavioral ecology; it only counts as evo psych if the method is ropey and the evidence sparse”.”
You can only say that if you do not actually known the literature in evopsych and human behavioral ecology. I do.
LikeLike